


流感嗜血杆菌是呼吸道常见定植菌,可继发于病毒感染、支原体感染后引起鼻窦炎、中耳炎、肺炎等呼吸道感染[1,2]。在儿童及免疫力低下的老年人群,流感嗜血杆菌也可引起菌血症、脑膜炎、关节脓肿或其他部位的侵袭性感染[3]。根据有无荚膜,流感嗜血杆菌分为有荚膜和无荚膜的菌株即不可分型的流感嗜血杆菌(NTHi)[3]。有荚膜的流感嗜血杆菌多引起全身感染,可以分为a ~ f共6种血清型。其中,b型流感嗜血杆菌(Hib)是引起会厌炎、脑膜炎及菌血症的主要亚型[4]。无荚膜的菌株多引起上呼吸道部位的感染,但在儿童及免疫低下的老人可导致会厌炎、乳突炎和肺炎[5]。
氨苄西林、头孢呋辛、头孢噻肟是治疗流感嗜血杆菌感染的一线药物。β-内酰胺类抗菌药物的广泛使用使流感嗜血杆菌耐药率不断升高,引起了临床的广泛关注[6]。据报道,流感嗜血杆菌的耐药机制主要是携带TEM型和ROB型β-内酰胺酶[1]。
本研究对分离的流感嗜血杆菌进行药物敏感性、耐药基因检测及血清型分析,明确济南地区流感嗜血杆菌的分子流行病学特征,比较不同基因型菌株对抗菌药物的耐药情况,为经验性治疗流感嗜血杆菌感染提供依据。
材料与方法
一、材 料
1. 菌株来源
收集2018年1至10月在山东大学附属省立医院住院患者的送检标本,培养分离流感嗜血杆菌,剔除来自同一患者同一感染部位的重复分离菌株。采用梅里埃质谱鉴定仪对流感嗜血杆菌进行菌种鉴定。质控菌株流感嗜血杆菌ATCC 49247、粪肠球菌ATCC 29212、金黄色葡萄球菌ATCC 29213均为本实验室保存。
2. 主要仪器及试剂
时间飞行质谱仪(MALDI-TOF MS,法国梅里埃公司),细菌浊度仪(法国梅里埃公司),恒温细菌培养箱(上海精宏实验设备有限公司),PCR扩增仪(美国应用生物系统公司),Bio-Rad核酸电泳仪 (美国伯乐公司),GSG-2000凝胶成像系统(中国黑马医学仪器)。头孢硝噻吩纸片( 重庆庞统医疗) ;流感嗜血杆菌药物敏感性试剂盒(温州康泰公司)。引物委托上海生物工程有限公司合成,PCR Premix Taq™ 购自大连宝生物有限公司,Gel-Red核酸染料、50×TBE 电泳缓冲液、100 bp DNA Maker、核酸琼脂糖凝胶为百泰克公司产品。
二、方 法
1. 药物敏感试验及数据分析
参照CLSI M100 S28,采用微量肉汤稀释法对流感嗜血杆菌进行抗菌药物的药物敏感试验和结果判读。药物敏感质控菌株为流感嗜血杆菌ATCC 49247。
2. β-内酰胺酶表型检测
双蒸水浸湿纸片后沾取待检测菌苔,室温放置1 h观察结果。粪肠球菌ATCC 29212和金黄色葡萄球菌ATCC 29213分别作为β-内酰胺酶阴性和阳性对照。
3. 耐药基因PCR检测
3.1 引物合成
表1 PCR扩增引物序列 |
| 基因名称 | 引物序列 | 产物长度(bp) | 退火温度 |
|---|---|---|---|
| TEM | 正向:5’-TGGGTGCACGAGTGGGTTAC-3’ | 526 | 57℃ |
| 反向:5’-TTATCCGCCTCCATCCAGTC-3’ | |||
| ROB | 正向:5’-ATCAGCCACACAAGCCACCT-3’ | 692 | 62℃ |
| 反向:5’-GTTTGCGATTTGGTATGCGA-3’ | |||
| bexA | 正向:5’-CGTTTGTATGATGTTGATCCAGAC-3’ | 567 | 54℃ |
| 反向:5’-TGTCCATGTCTTCAAAATGATG-3’ | |||
| bexB | 正向:5’-GGTGATTAACGCGTTGCTTATGCG-3’ | 343 | 54℃ |
| 反向:5’-TTGTGCCTGTGCTGGAAGGTTATG-3’ | |||
| a | 正向:5’-CTACTCATTGCAGCATTTGC-3’ | 250 | 60℃ |
| 反向:5’-GAATATGACCTGATCTTCTG-3’ | |||
| b | 正向:5’-GCGAAAGTGAACTCTTATCTCTC-3’ | 480 | 60℃ |
| 反向:5’-GCTTACGCTTCTATCTCGGTGAA-3’ | |||
| c | 正向:5’-TCTGTGTAGATGATGGTTCA-3’ | 250 | 60℃ |
| 反向:5’-CAGAGGCAAGCTATTAGTGA-3’ | |||
| d | 正向:5’-TGATGACCGATACAACCTGT-3’ | 150 | 60℃ |
| 反向:5’-TCCACTCTTCAAACCATTCT-3’ | |||
| e | 正向:5’-GGTAACGAATGTAGTGGTAG-3’ | 1350 | 60℃ |
| 反向:5’-GCTTTACTGTATAAGTCTAG-3’ | |||
| f | 正向:5’-GCTACTATCAAGTCCAAATC-3’ | 450 | 60℃ |
| 反向:5’-CGCAATTATGGAAGAAAGCT-3’ |
3.2 模板制备
取适量流感嗜血杆菌菌苔充分混匀于500 μl去离子水中,浓度约为1.00 D。沸水浴中煮10 min,12 000×g离心10 min,将上清液转移至干净的EP管中即为DNA模板,保存于-20℃冰箱备用。
3.3 PCR反应体系
引物各0.2 μmol/L,PCR Premix 10 μl,DNA模板2 μl,双蒸水补齐至20 μl[10]。
3.4 耐药基因扩增条件
TEM基因PCR扩增条件为:94℃预变性10 min;94℃变性1 min,57℃退火45 s,72℃延伸1 min,共30个循环;72℃延伸10 min。ROB基因扩增条件为:94℃预变性2 min;94℃变性2 min,62℃退火45 s,72℃延伸2 min,共30个循环;72℃终延伸10 min[9]。
3.5 荚膜分型扩增条件
bexA基因扩增条件为:94℃预变性2 min;94℃变性2 min,54℃退火30 s,72℃延伸2 min,共30个循环;72℃终延伸10 min。bexB基因扩增条件为:94℃预变性2 min;94℃变性2 min,54℃退火45 s,72℃延伸2 min,共30个循环;72℃终延伸10 min。a ~ f群分型扩增条件为:94℃预变性2 min;94℃变性2 min,60℃退火45 s,72℃延伸2 min,共30个循环;72℃终延伸10 min[7]。
3.6 PCR产物电泳
采用1.5%的琼脂糖凝胶,0.5×TBE缓冲液,5 μl PCR产物上样,110 V电泳电压,电泳时间为40 min。黑马成像系统紫外灯下观察条带并拍照。阳性样本送交上海生物工程有限公司测序确认。
三、统计学处理
采用WHONET 5.6 进行药物敏感性分析,并对成年人与未成年人分离株耐药率(不含中介率),以及产酶株与不产酶株耐药率(不含中介率)进行了比较。应用SPSS 13.0进行统计分析,组间比较采用$\chi$2检验、校正$\chi$2检验或Fisher确切概率法,P < 0.05为差异有统计学意义。
结 果
一、菌株标本来源
107株流感嗜血杆菌分离自痰液94株(87.85%),鼻腔脓液10株(9.30%),肺泡灌洗液3株(2.80%)。41株(38.32%)标本来自未成年人(年龄小于18岁)患者,66株(61.68%)标本来自成人(年龄 ≥18岁)患者。
二、流感嗜血杆菌抗菌药物耐药性
临床共分离107株流感嗜血杆菌,菌株对于各药物的耐药率见表2。其中,美罗培南的药物敏感率为100%,其次为头孢噻肟和复方磺胺甲噁唑,细菌敏感率大于90%。
表2 107株流感嗜血杆菌对8种抗菌药物耐药性 株数(%) |
| 抗菌药物 | 耐药 | 中介 | 敏感 |
|---|---|---|---|
| 氨苄西林/舒巴坦 | 49(45.8) | 1(0.9) | 57(53.3) |
| 氯霉素 | 11(10.3) | 4(3.7) | 92(86.0) |
| 氨苄西林 | 82(76.6) | 4(3.7) | 21(19.6) |
| 美罗培南 | 0 | 0 | 107(100) |
| 头孢噻肟 | 10(9.3)a | 0 | 97(90.7) |
| 头孢呋辛 | 44(41.1) | 5(4.7) | 58(54.2) |
| 复方磺胺甲 口 恶 唑 | 73(68.2) | 2(1.9) | 32(29.9) |
| 左氧氟沙星 | 5(4.7)a | 0 | 102(95.3) |
注:a为非敏感率 |
三、成人与未成年人流感嗜血杆菌的耐药性比较
成人组中氨苄西林/舒巴坦、氯霉素、头孢呋辛耐药率明显高于未成年人组;未成年人组中氨苄西林、头孢噻肟、复方复方磺胺甲噁唑、左氧氟沙星的耐药率高于成人组。未成年人组与成人组药物敏感性比较见表3。
表3 成人与未成年人流感嗜血杆菌对8种抗菌药物耐药性比较 |
| 抗菌药物 | 耐药率(%) | $\chi^{2}$值 | P值 | |
|---|---|---|---|---|
| 未成年人组 (n = 41) | 成人组 (n = 66) | |||
| 氨苄西林/舒巴坦 | 43.9 | 47.0 | 0.100 | 0.760 |
| 氯霉素 | 4.9 | 13.6 | 0.479b | 0.489 |
| 氨苄西林 | 85.4 | 71.2 | 2.830 | 0.090 |
| 美罗培南 | 0 | 0 | - | - |
| 头孢噻肟 | 2.4a | 1.5a | 0.178b | 0.673 |
| 头孢呋辛 | 34.2 | 45.5 | 1.340 | 0.250 |
| 复方磺胺甲 口 恶 唑 | 73.2 | 65.2 | 0.750 | 0.390 |
| 左氧氟沙星 | 2.4a | 1.5 | 0.178b | 0.673 |
注:a为非敏感率;b校正$\chi$2检验 |
四、产酶株与不产酶株药物耐药性比较
107株流感嗜血杆菌β-内酰胺酶阳性菌株为72株(67.3%),全部表现为对氨苄西林耐药。在产β-内酰胺酶菌株中约55.6%对氨苄西林/舒巴坦耐药。产酶株对氨苄西林、氨苄西林/舒巴坦耐药率高于非产酶株,差异有统计学意义。产酶株与不产酶株药物敏感性比较见表4。
表4 产酶株与不产酶株药物耐药性比较 |
| 抗菌药物 | 耐药率(%) | $\chi$2值 | P值 | |
|---|---|---|---|---|
| β-内酰胺酶(+) (n = 72) | β-内酰胺酶(-) (n = 35) | |||
| 氨苄西林/舒巴坦 | 55.6 | 20 | 12.090 | <0.001 |
| 氯霉素 | 11.1 | 8.6 | 0.004b | 0.947 |
| 氨苄西林 | 100 | 22.9 | 74.290 | <0.001 |
| 美罗培南 | 0 | 0 | - | - |
| 头孢噻肟 | 1.4a | 2.9a | - | 1.000c |
| 头孢呋辛 | 43.1 | 31.4 | 1.340 | 0.250 |
| 复方磺胺甲 口 恶 唑 | 69.4 | 60 | 0.940 | 0.330 |
| 左氧氟沙星 | 2.78a | 0 | - | 1.000c |
注:a为非敏感率,b校正$\chi$2检验,c Fisher确切概率法 |
五、耐药基因检测
107株流感嗜血杆菌菌株中,共72株检测到β-内酰胺酶基因(69.2%),与头孢硝噻吩纸片检测β-内酰胺酶结果一致。产酶株中70株携带TEM型基因,2株携带ROB基因,见图1。
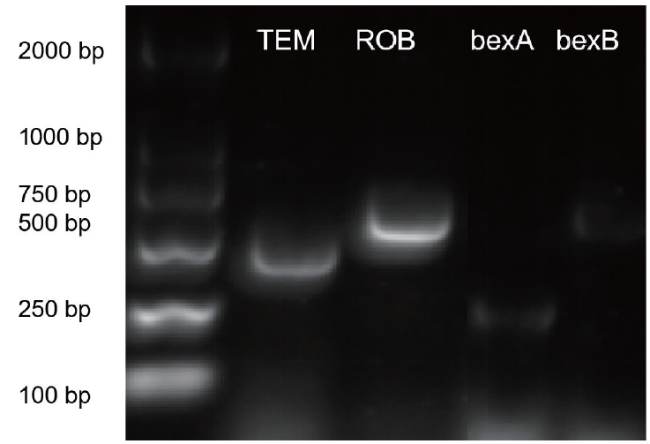
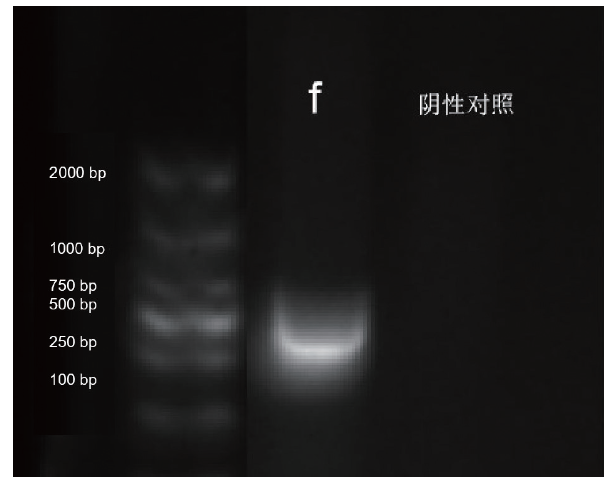
六、流感嗜血杆菌荚膜分型
{kind=link}
{kind=link}
{kind=link}
{kind=link}
讨 论
本次研究中107株流感嗜血杆菌均分离自呼吸道标本,是引起呼吸道感染的重要病原菌。本研究中,流感嗜血杆菌对氯霉素、美罗培南、头孢噻肟、左氧氟沙星的敏感率较高,均超过85%;氨苄西林的敏感率仅为19.6%。与2017年全国细菌耐药监测网儿童及新生儿患者数据进行比较,本地区氨苄西林(76.6% vs. 62.9%)、氨苄西林/舒巴坦(45.8% vs. 30.7%)、头孢呋辛(41.1% vs. 36.1%)、左氧氟沙星(4.7% vs. 0.8%)耐药率或不敏感率均高于全国平均水平,其他药物敏感性与全国平均水平基本一致[2]。β-内酰胺类及β-内酰胺类/β-内酰胺酶抑制剂类药物耐药率的升高可能与本地区用药习惯有关,应引起关注。流感嗜血杆菌目前对于三代头孢菌素仍有较好敏感性,可以供临床经验性用药选择。
流感嗜血杆菌对氨苄西林耐药于1972年首先在欧洲报道,主要耐药机制是产生β-内酰胺酶,编码β-内酰胺酶的基因主要存在于质粒DNA上,少部分位于染色体DNA上[9, 11]。本研究中,107株流感嗜血杆菌β-内酰胺酶检出率为67.3%,远高于文献中54.8%的比例。产β-内酰胺酶(TEM型或ROB型)是流感嗜血杆菌对β-内酰胺酶类耐药的主要机制。此次对107株临床分离株进行检测发现72株产TEM型β-内酰胺酶,2株产ROB型β-内酰胺酶。107株流感嗜血杆菌中共10株(9.3%)为β-内酰胺酶阴性氨苄西林耐药(BLNAR),该比例与国内报道基本一致。BLNAR的耐药机制不同于产酶机制,为编码细胞壁的青霉素结合蛋白3的ftsI基因点突变导致[5]。该点突变除了引起青霉素的耐药,还可引起其他β-内酰胺酶药物治疗的失败。近年来,BLNAR的升高已经引起临床的重视。
本研究中,只有一株流感嗜血杆菌可以划分为血清型f型,其余106株菌株均为NTHi,与田磊等[3]的报道一致。这可能与本地区Hib疫苗接种及本研究入组人群以成人为主有关。据报道,NTHi感染率逐渐上升,已成为流感嗜血杆菌的主要致病血清型,与其表面表达多种黏附素有关。
综上所述,流感嗜血杆菌对氨苄西林耐药率逐渐升高,不建议该药继续作为临床治疗流感嗜血杆菌感染的首选药物。BLNAR及NTHi的发生率也在悄然升高,应引起关注。大规模流感嗜血杆菌分子流行病学调查及新型疫苗的研发有待开展。