


肝细胞癌(HCC)是全球最常见的恶性肿瘤之一,也是导致癌症死亡的第二大原因,占原发性肝癌的80%,严重威胁人类健康[1,2]。随着对HCC分子生物学研究的不断深入,已鉴定出与该疾病有关的不同基因和途径。为了更好地理解HCC发生、发展的分子机制,亟待发现更精准的预后标志物及有效的治疗策略。张春艳等(2016年)认为作为一类非编码单链RNA分子,微RNA(miR)与肝癌的发生、发展过程密切相关。miR-144的高表达可明显抑制裸鼠肝癌形成[3]。近期研究表明,miR-190可以通过抑制ZEB2的表达来抑制胶质瘤细胞的生长和迁移[4]。然而,miR-190在HCC中的表达、潜在作用和机制仍不清楚。本研究探讨miR-190通过调控细胞因子信号转导抑制因子5(SOCS5)参与HCC发生与发展过程的关键作用与机制。
材料与方法
一、研究对象
选取2019年5月至2019年12月在上海中医药大学附属曙光医院接受手术切除且病理确诊为HCC的患者84例,其中男58例、女26例,年龄(63±5)岁。收集HCC癌组织及对应的癌旁组织(距肿瘤边缘≥2 cm)。患者均签署知情同意书,研究方案经上海中医药大学附属曙光医院伦理委员会审核通过。
二、细胞、动物及主要试剂
SNU398和HepG2细胞均购于中国科学院上海细胞研究所。24只雌性无胸腺裸鼠,4 ~ 6周龄,购自中国医学科学院实验动物中心。SOCS5、β-actin抗体购自Sigma公司;miR-190模拟物/抑制物和Dharmafect 1购自Dharmacon公司;靶向SOCS5的psiCHECK-2载体购自上海联迈生物工程有限公司;Lipofectamine 3000转染试剂、TRIzol试剂、miR定量试剂盒购自Thermo Fisher Scientific公司;Transwell小室购自Corning公司;Mgteigel基质胶购自BD公司。
三、细胞培养与转染
SNU398和HepG2细胞分别用含10%胎牛血清、100 U/ml青霉素、100 μg/ml链霉素的RPMI 1640或DMEM培养基于5% CO2、37°C恒温培养箱中培养。在转染前将细胞接种于12孔板中,待细胞达到每孔150 000个细胞的密度时,根据说明书将miR-190模拟物/抑制物或SOCS5 siRNA(siSOCS5)及各自的阴性对照(模拟物NC/抑制物NC或siNC)分别用Dharmafect 1和Lipofectamine 3000转染至细胞。
四、细胞活力
根据细胞计数试剂盒(CCK-8)说明书测定细胞活力。将转染细胞按1×103 个/孔接种于96孔板,每孔加入10 μl CCK-8 溶液,在37℃下孵育2 h。用酶标仪检测450 nm波长下细胞的吸光度。
五、Transwell侵袭与迁移实验
取100 μl基质胶稀释液铺于Transwell小室上室,含胎牛血清的常规培养基加入下室,细胞置于37 ℃、5% CO2 培养箱内继续培养24 h。擦去上室细胞与基质胶,4%多聚甲醛固定,1%结晶紫染色,显微镜下观察并计算侵袭细胞数。迁移实验无需铺胶,其余操作同侵袭实验。
六、裸鼠异种移植肿瘤
于裸鼠腹侧皮下接种1.5×106个用siNC或siSOCS5转染的SNU398或HepG2细胞。对肿瘤生长进行长达18 d的监测,并通过以下公式计算肿瘤体积:体积 = (L×W2)/2,其中L为肿瘤的长度,W为肿瘤的宽度。
七、RNA提取和实时荧光定量PCR(RT-PCR)
采用TRIzol法提取HCC组织、癌旁组织、转染后的SNU398或HepG2细胞中的总RNA。根据miR逆转录试剂盒说明书进行逆转录,随后,使用7900HT快速实时PCR系统(Thermo Fisher Scientific)进行RT-PCR。miR-190以U6为内参,SOCS5以β-actin为内参,采用2-ΔΔCt方法计算结果。引物序列见表1。
表1 PCR引物序列 |
| 转录因子 | 上游引物 | 下游引物 |
|---|---|---|
| miR-190 | 5'-TGACTCCAAGTACTGGAGGT-3' | 5'-TAGCCGCCTATACGGAATGG-3' |
| U6 | 5'-GCGCGTCGTGAAGCGTTC-3' | 5'-GTGCAGGGTCCGAGGT-3' |
| SOCS5 | 5'-TGGAACAATAGCGGATAGAGC-3' | 5'-TTGCCTCACTCAAATACCACA-3' |
| β-actin | 5'-ACACCTTCTACAATGAGCTG-3' | 5'-CTGCTTGCTGATCCACATCT-3' |
八、蛋白免疫印迹法
利用RIPA裂解液提取细胞蛋白,并用Bradford法测定蛋白质浓度。将10 μg总蛋白上样后,进行SDS-PAGE电泳,并转移至PVDF膜上。封闭2 h后,加入SOCS5抗体(1∶1000)或β-actin抗体(1∶2000),4℃孵育过夜。TBST冲洗3次后,加入二抗,室温孵育l h,洗膜后用ECL化学发光试剂盒显影。
九、荧光素酶测定
构建野生型和突变型基因靶点SOCS5的3′UTR-荧光素酶表达载体(SOCS5-Wt和SOCS5-Mut),使用Lipofectamine 3000将其与miR-190模拟物和阴性对照同时分别转染。转染72 h后,使用双荧光素酶报告系统(上海吉马生物制药有限公司)连续测量萤火虫-海肾荧光素酶的活性。
十、统计学处理
应用GraphPad Prism 8.0分析数据,正态分布定量资料以 $\bar{x} \pm s$表示,定性资料以例表示。采用配对t检验分析癌组织和癌旁组织间的差异,独立样本t检验分析体外定量数据,两因素重复测量方差分析转染时间与分组间的差异,x2检验分析miR-190表达与临床特征的关系。生存曲线采用Kaplan-Meier 法,log-rank检验进行比较。α = 0.05。
结果
一、miR-190是HCC中潜在的癌基因
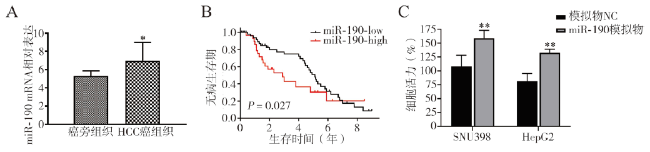
图1 miR-190表达与HCC的关系A:HCC癌组织和及癌旁组织miR-190的mRNA表达水平;B:miR-190表达水平高低与HCC患者预后的相关性;C:SNU398和HepG2细胞的miR-190的表达对细胞体外增殖能力的影响;*P < 0.05;**P < 0.01 |
表2 miR-190表达量与HCC患者临床特征的关系[例(%)] |
| 临床特征 | miR-190高表达(40例) | miR-190 低表达 (44例) | x2值 | P值 |
|---|---|---|---|---|
| 年龄 | 0.119 | 0.730 | ||
| ≤60岁 | 14(35) | 17(39) | ||
| >60岁 | 26(65) | 27(61) | ||
| 性别 | 0.585 | 0.444 | ||
| 男 | 26(65) | 32(73) | ||
| 女 | 14(35) | 12(27) | ||
| 肿瘤大小 | 5.002 | 0.025 | ||
| ≤5 cm | 13(32) | 25(57) | ||
| >5 cm | 27(68) | 19(43) | ||
| 肿瘤个数 | 0.087 | 0.768 | ||
| 1个 | 24(60) | 25(57) | ||
| ≥2个 | 16(40) | 19(43) | ||
| 淋巴结转移 | 8.091 | 0.004 | ||
| N0 | 20(50) | 35(80) | ||
| N+ | 20(50) | 9(20) | ||
| 临床分期 | 10.309 | 0.001 | ||
| Ⅰ~Ⅱ | 26(65) | 41(93) | ||
| Ⅲ~Ⅳ | 14(35) | 3(7) |
二、下调miR-190的表达抑制肝癌细胞侵袭及迁移能力
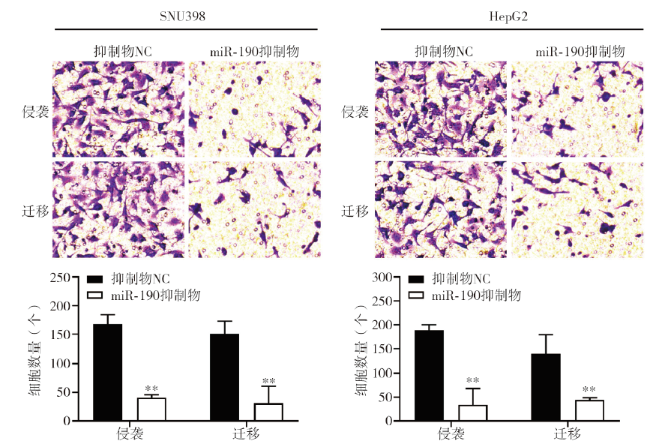
下调miR-190处理SNU398和HepG2肝癌细胞后,发现下调miR-190的表达抑制了肝癌细胞的侵袭(t = 13.120, P < 0.001;t = 7.398, P = 0.002)及迁移能力(t = 5.587, P = 0.005;t = 4.082,P = 0.015),见图2。
三、miR-190直接靶向SOCS5
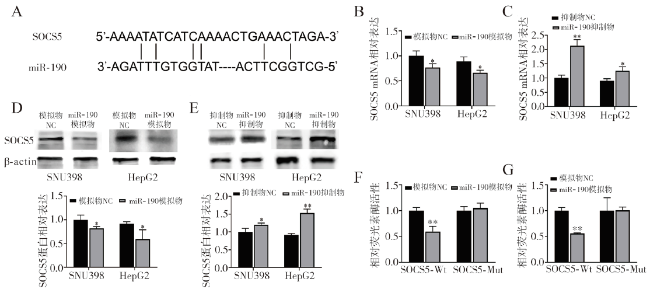
借助生物信息学数据库Star Base 3.0对miR-190的靶基因进行预测,发现SOCS5可能是miR-190的潜在靶点(图3A)。为了验证该假设,我们通过RT-PCR和蛋白免疫印迹法检测了miR-190对SOCS5转录和蛋白质水平的影响。结果显示,miR-190的过表达导致SNU398和HepG2细胞中SOCS5 mRNA(t = 3.212, P = 0.033;t = 3.597, P = 0.023, 图3B)和蛋白(t = 2.895, P = 0.044;t = 2.813, P = 0.048,图3D)表达减少,而抑制miR-190导致SOCS5 mRNA(t = 8.249, P = 0.001;t = 3.865, P = 0.018, 图3C)和蛋白表达(t = 3.098, P = 0.036;t = 8.835, P = 0.001,图3E)的增加。
四、SOCS5是HCC的肿瘤抑制因子
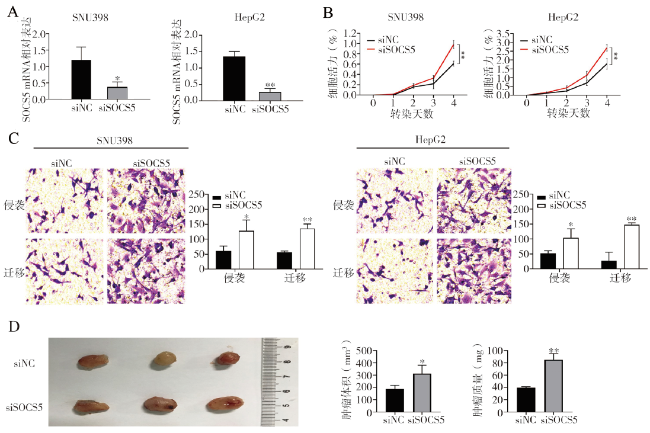
靶向siRNA可有效下调SNU398和HepG2细胞中的SOCS5水平(t = 3.342, P = 0.028;t = 9.887, P = 0.001, 图4A)。
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
在体外,两因素重复测量方差分析显示SNU398和HepG2细胞的增殖随时间变化呈上升趋势(F时间 = 277.7和233.3,P均< 0.001,图4B),siSOCS5组增殖能力高于siNC组(F组间 = 36.6和34.2,P均< 0.001),分组与时间有交互效应(F = 15.4和9.656, P均< 0.001)。此外,转染siSOCS5的细胞迁移(t = 2.946, P = 0.042;t = 7.258, P = 0.002)和侵袭能力(t = 2.876, P = 0.045;t = 8.764, P = 0.001)均增强(图4C)。我们在异种移植肿瘤模型中进一步评估了SOCS5基因敲除对HCC细胞肿瘤发展过程中的作用。与阴性对照相比,SOCS5基因敲除导致SNU398和HepG2细胞的异种移植肿瘤体积(t = 2.829, P = 0.047)和质量(t = 7.635, P = 0.002)增加(图4D)。
讨论
有研究显示在侵袭性神经母细胞瘤和前列腺癌中miR-190表达减少,而在快速生长的肿瘤中其过度表达会抑制肿瘤生长和转移,miR-190还调节上皮间质转化抑制乳腺癌转移[5]。相反,Jia等(2016年)研究显示miR-190在胃癌组织中表达上调,并促进胃癌的进展。这提示miR-190可能在不同的肿瘤环境和肿瘤发展的不同阶段发挥不同的作用。而关于miR-190在肝癌中的作用的研究目前较少,如Hung等(2014年)发现miR-190b的上调对诱导人HCC胰岛素抵抗的IGF-1降低起作用。本研究首先发现miR-190在HCC肿瘤中上调,且miR-190高表达与HCC患者的不良预后有关,提示其可能在HCC发展过程中起促进作用。此外,我们检测了miR-190过表达对HCC细胞生长的影响,结果发现miR-190过表达能促进细胞增殖,而下调miR-190抑制了肝癌细胞的侵袭及迁移能力,表明miR-190通过促进HCC细胞增殖、侵袭和迁移等来发挥其促癌作用。这些结果与在胃癌中的观察结果相一致。
当miR-190过表达时,SOCS5在HCC细胞系中下调,由此我们推测SOCS5是miR-190的潜在靶点。SOCS5在人类癌症中的作用已被研究,在前列腺癌和肝癌中已经检测到SOCS5的下调[6]。据报道,抑制miR-18a-5p促进SOCS5而诱导骨肉瘤细胞凋亡[7]。随后,我们使用荧光素酶检测法证明了它确实是miR-190的靶点,同时,miR-190还能调节其转录和蛋白水平。有趣的是,SOCS基因是JAK/STAT通路中负反馈回路的一部分[8]。众所周知,JAK/STAT途径可激活细胞的增殖、迁移、分化、凋亡以及抑制剂的失调,导致包括肝癌在内的人类疾病[9]。Yuan等[10]研究发现,miR-30a靶向SOCS-1,并通过JAK/STAT信号通路抑制脓毒症大鼠的肝细胞增殖和促进细胞凋亡。Wang等[11]发现了板蓝根多糖通过激活JAK/STAT信号通路来发挥对HBV的体外抗病毒作用。考虑到目前SOCS5与肝癌的相关研究较少,我们进行了额外实验来验证SOCS5在肝癌中的作用,结果证明了SOCS5在体内和体外都可作为HCC的肿瘤抑制因子。以上结果表明,在HCC中,由miR-190介导的SOCS5下调将导致JAK/STAT通路的激活,进而促进细胞增殖、侵袭和迁移。有研究报道miR-885-5p上调通过靶向抑制SOCS5促进大肠癌细胞增殖和迁移,我们的研究结果与其一致[12]。
综上所述,我们验证了细胞因子家族抑制因子之一的SOCS5作为miR-190在肝癌中的真实靶点,并证明了SOCS5在肝癌中的抑瘤作用。此外,我们强调了miR190的抑制作用在恢复SOCS5水平方面的潜在用途,从而降低肝癌细胞的致瘤特性。这项研究强调了miR-190在肝癌研究中的重要性,不仅是作为潜在的生物标志物,而且有可能成为待开发潜在药物的新的分子靶标。