


肥胖是全球重大的公共健康问题[1]。肥胖改变全身代谢并导致胰岛素抵抗,其机制涉及脂肪组织的线粒体功能损伤和炎症[2]。间歇性禁食(IF)是指在特定时间段内限定热量摄入的一种饮食管理手段[3]。研究表明,IF可以作为一种有效的行为干预方式来减少包括肥胖在内的代谢性疾病的负担及延长寿命,但具体机制尚未完全明确[4]。沉默信息调节因子2相关酶1(SIRT1)是一种依赖烟酰胺腺嘌呤二核苷酸(NAD+)的去乙酰化酶,表达受能量状态调节,NAD+高表达可改善高脂饮食诱导的肥胖和胰岛素抵抗形成,并改善脂肪组织和肝脏的线粒体功能,减轻氧化应激和炎症程度[5⇓⇓⇓⇓-10]。因此推测,SIRT1在IF改善肥胖中可能发挥重要作用。本研究通过构建隔日禁食的间歇性高脂饮食小鼠模型、利用尾静脉注射腺相关病毒(AAV)敲减SIRT1等手段,探讨IF对高脂诱导的肥胖小鼠脂肪组织线粒体功能、炎症通路的影响以及SIRT1在其中的介导作用,为揭示SIRT1在IF调控机体代谢平衡中的作用提供理论基础。
材料与方法
一、动物模型的建立及IF方案
5~6周龄雄性C57/BL6小鼠35只,体重18~22 g,购至江苏集萃药康生物科技股份有限公司。动物饲养于中山大学SPF级环境中,控制温度、湿度,进行12 h光-暗循环。小鼠经过1周适应性喂养后,将15只小鼠根据空腹血糖和体重随机分为普通自由饮食组(CD组,n = 5)、高脂自由饮食组(HFD组,n = 5)和高脂IF组(HFD-IF组,n = 5),HFD-IF组采取隔日禁食24 h、自由饮水,喂养12周。第2批小鼠适应性喂养1周后,将16只小鼠随机分为HFD组和HFD-IF组,干预4周后,通过尾静脉分别注射腺相关敲减SIRT1病毒和腺相关对照病毒,分别设为高脂自由饮食对照组(shCtrl-HFD组,n = 4)、高脂自由饮食敲减组(shSIRT1-HFD组,n = 4)、高脂IF对照组(shCtrl-HFD-IF组,n = 4)、高脂IF敲减组(shSIRT1-HFD-IF组,n = 4),继续喂养8周。所有小鼠实验结束后,用异氟烷麻醉后摘除眼球取血,采用颈椎脱臼法将其处死,留取小鼠附睾旁脂肪组织,切取部分组织置于4%多聚甲醛固定,将其余组织储存于-80℃冰箱。本实验通过中山大学实验动物伦理委员会审批(批件号:SYSU-IACUC-2021-000374)。
二、实验试剂和材料
1.主要试剂
高脂饲料(货号D12492)购自Research Diets公司(美国);AAV-shRNA-SIRT1和AAV-shRNA-Control购自广州东泽生物科技有限公司(中国),SIRT1干扰序列5′-TCGAACAATTCTTAAAGAT-3′,RIPA裂解液、蛋白酶抑制剂和BCA试剂盒购自Thermo Fisher公司(美国);磷酸酶抑制剂(货号CW2383S)购自康为世纪(中国);TRIzol试剂(货号T9424)购自Sigma-Aldrich公司(美国);Prime ScriptTMRT Master Mix逆转录试剂盒(货号RR036A)购自TaKaRa公司(日本);LightCycler 480 SYBR Green I Master购自Roche公司(美国);SIRT1、腺苷酸激活蛋白激酶(AMPK)、p-AMPK、叉头转录因子O1(FOXO1)、β-actin抗体均购自Cell Signaling Technology公司(美国);IRDye 800CW二抗(货号926-32211)购自LI-COR公司(美国);甘油三酯(货号A110-1-1)、总胆固醇试剂盒(货号A111-1-1)购自南京建成生物工程研究所(中国江苏)。
2. 主要仪器
荧光倒置显微镜及体视显微镜(Leica);Nanadrop2000分光光度计(Thermo);实时荧光定量PCR仪(Roche LightCycler 480);全自动酶标仪(Thermo);双色红外成像系统Odyssey(LI-COR)。
三、方法
1. HE染色
将小鼠脂肪组织固定,经过浸蜡、包埋、切片后,按照步骤进行二甲苯脱蜡,苏木素和0.5%伊红分别染色、脱水和封片,显微镜下观察并拍照。采用Image J软件统计脂滴大小。
2.组织蛋白提取和蛋白免疫印迹
取小鼠脂肪组织100 mg~200 μL预冷RIPA裂解液中,匀浆后采用14 000转/分于4℃离心20 min,吸取中间清液层得到蛋白原液。用BCA试剂盒检测蛋白浓度配平后,取30 μg总蛋白进行凝胶电泳,将蛋白转移到聚偏二氟乙烯膜上,用5%脱脂牛奶室温封闭1 h后,加入相应的Ⅰ抗稀释液(1∶1 000)于4℃过夜孵育。孵育结束后用TBST洗膜3次,加入Ⅱ抗稀释液(1∶10 000)室温避光孵育1 h后,再次洗膜,使用Odyssey红外荧光成像系统扫描,并用Image J软件测量蛋白条带灰度值进行定量。
3. 组织RNA提取和RT-qPCR检测
使用TRIzol试剂提取小鼠脂肪组织总RNA,用分光光度计测定总RNA浓度和纯度,用逆转录试剂盒将RNA逆转录为cDNA。使用试剂盒并采用PCR检测仪进行PCR扩增。反应体系的配制及反应程序均按照试剂盒说明书。内参照为β-actin,结果使用2-∆∆Ct方法分析计算目的基因的相对表达量。
4.血清检测
小鼠血清中甘油三酯(TG)和总胆固醇(TC)水平的测定按照相应试剂盒进行测定。
四、统计学处理
采用GraphPad Prism 8.0进行统计分析和图表绘制分析。数据采用$\bar{x}\pm s$表示。数据符合正态分布,多组比较采用单因素方差分析,进一步两两比较采用LSD-t检验;P < 0.05为差异有统计学意义。
结果
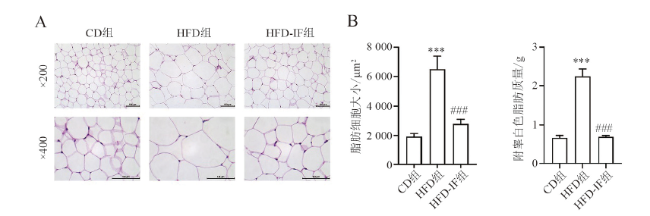
一、IF能减小高脂饮食喂养小鼠脂肪细胞体积并降低血脂水平
小鼠脂肪组织HE染色和称重结果显示,与CD组相比,HFD组小鼠脂肪细胞体积增大且质量更重(F = 30.732,P < 0.001;F = 63.267,P < 0.001),而HFD-IF组小鼠脂肪细胞体积较HFD组减小且质量减轻(F = 30.732,P < 0.001;F =63.267,P < 0.001),脂质含量明显降低,见图1。HFD组小鼠血清中TG、TC水平高于CD组[TG HFD vs. CD:(148.66±11.44)mg/dL vs.(79.33±5.43) mg/dL,F = 22.913, P < 0.001;TC HFD vs. CD:(160.40±7.62) mg/dL vs. (100.12±2.05) mg/dL,F = 38.717, P < 0.001],而HFD-IF组较HFD组小鼠水平下降[TG HFD-IF vs. HFD:(80.96±6.69)mg/dL vs. (148.66±11.44)mg/dL,P < 0.001;TC HFD-IF vs. HFD:(116.23±3.63)mg/dL vs. (160.40±7.62)mg/dL,P < 0.001]。
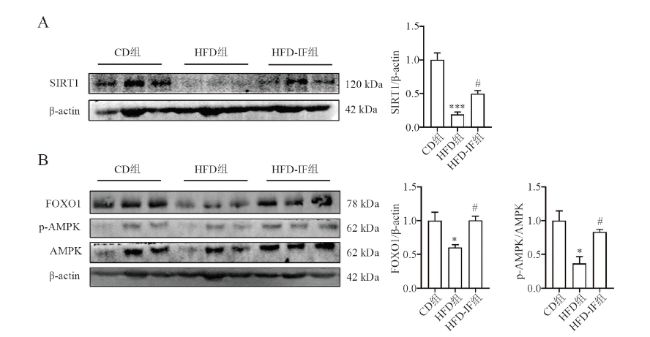
二、IF上调高脂饮食小鼠白色脂肪组织中SIRT1及激活AMPK、FOXO1通路
蛋白免疫印迹结果显示,HFD组小鼠中SIRT1、p-AMPK、FOXO1表达均较CD组降低(F = 35.641,P < 0.001;F = 7.503,P = 0.037;F =9.935,P = 0.012),而IF使SIRT1、p-AMPK、FOXO1水平升高(F = 35.641,P = 0.043;F = 7.503,P = 0.034;F = 9.935、P = 0.044)。见图2。
三、IF增加高脂饮食小鼠白色脂肪组织线粒体生物合成并减轻炎症程度
与CD组相比,HFD组小鼠脂肪组织中线粒体生物合成相关基因Pgc-1a、Tfam和Nrf1的mRNA相对表达量均降低(F = 23.561,P < 0.001;F = 15.872,P < 0.001;F = 24.456,P < 0.001),而炎症相关基因TNF-α、单核细胞趋化蛋白1(MCP-1)和生长因子样模体黏液样激素样受体(F4/80)的mRNA表达水平均升高(F = 16.742,P = 0.006;F = 17.743,P = 0.003;F = 8.492,P = 0.007);与HFD组相比,HFD-IF组小鼠线粒体生物合成相关基因表达水平上升(Pgc-1a、Tfam和Nrf1分别为P = 0.003、P = 0.022、P = 0.021),炎症相关基因表达下降(TNF-α、MCP-1和F4/80分别为P < 0.001、P < 0.001、P = 0.043)。
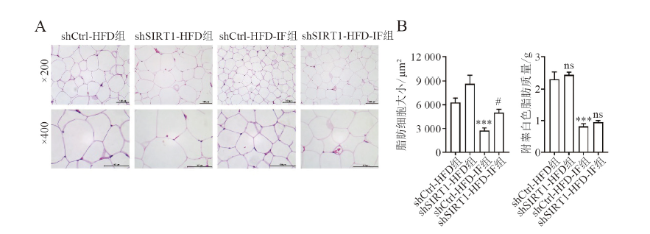
四、敲减SIRT1削弱IF减小高脂饮食喂养小鼠脂肪细胞体积和降低血脂的作用
通过尾静脉注射AAV-shSIRT1敲减SIRT1表达后,shSIRT1-HFD组与shCtrl-HFD组脂肪细胞体积大小比较差异无统计学意义(P > 0.05),而shSIRT1-HFD-IF组与对照组相比,脂肪质量无明显变化但细胞体积减小的程度有所下降(体积F = 17.993,P = 0.047;质量F = 46.439,P =0.881),见图3A~B。敲减SIRT1后,IF降低HFD诱导的高血脂作用被减弱(IF后TG shCtrl vs. shSIRT1:(82.30±2.12) mg/dL vs. (124.74±2.94) mg/dL,F = 97.605, P < 0.001;TC shCtrl vs. shSIRT1:(105.56±2.95) mg/dL vs. (144.83±5.10) mg/dL,F = 28.871, P = 0.002)。以上结果提示敲减SIRT1后,IF改善高脂饮食诱导的脂肪细胞体积增大和降低血脂的作用有所削弱。
五、敲减SIRT1减低间歇性禁食激活白色脂肪组织AMPK及FOXO1的作用
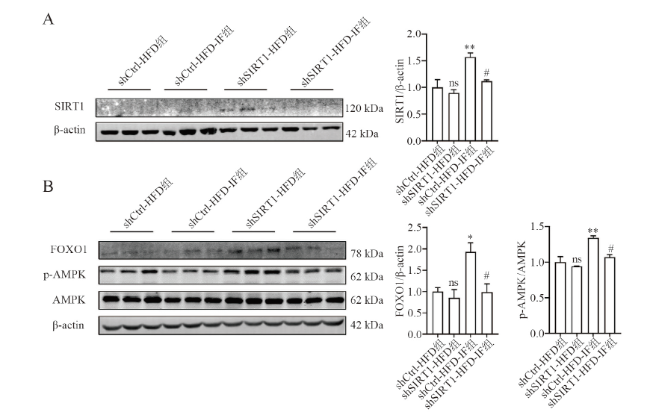
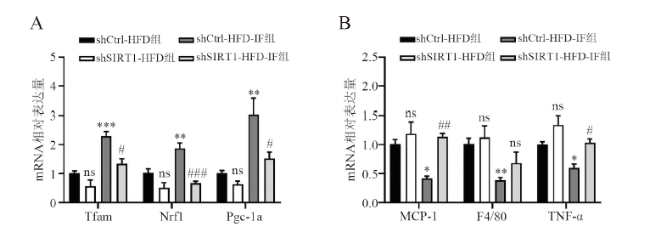
六、敲减SIRT1消除IF增加高脂饮食小鼠白色脂肪组织线粒体合成和炎症的作用
RT-qPCR结果显示,shCtrl-HFD-IF组与shCtrl-HFD组相比,线粒体功能相关基因如Tfam、Nrf1和Pgc-1a的mRNA表达均增加(F = 19.495,P < 0.001;F = 16.386,P = 0.008;F = 12.254,P =0.002),炎症相关基因MCP-1、F4/80和TNF-α的mRNA水平下调(F = 9.857,P = 0.014;F = 5.039,P = 0.050;F = 10.264,P = 0.047)。而敲减SIRT1后,与shCtrl-HFD-IF组相比,shCtrl-HFD-IF组IF的这种改善作用基本消失(Tfam P = 0.008、Nrf1 P < 0.001、Pgc-1a P = 0.018;MCP-1 P = 0.002、F4/80 P = 0.524、TNF-α P = 0.030)。见图5。
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
讨论
本课题组的前期研究证实SIRT1作为受能量代谢调控的去乙酰化蛋白,在高脂条件下其在肝脏中的表达降低,禁食时表达上调[15-16]。在本研究中,白色脂肪组织中SIRT1的蛋白表达结果与既往研究结果相似,相关研究证实,SIRT1可通过调控脂肪组织和肝脏的Pgc-1a促进线粒体生物合成,并能控制促炎基因转录从而调节脂肪组织炎症程度[9,17 -18]。本研究显示高脂诱导小鼠白色脂肪组织Pgc-1a、Tfam和Nrf1的表达降低,MCP-1、F4/80以及TNF-α的表达增加,提示存在线粒体功能障碍和脂肪炎症,IF能改善上述基因的表达;然而尾静脉注射AAV-shSIRT1敲低小鼠SIRT1表达后,IF对上述线粒体功能及炎症相关基因的作用被显著减弱,提示SIRT1可能通过改善线粒体功能障碍、减轻机体组织炎症程度而缓解机体代谢性疾病。
综上所述,IF可能通过调控脂肪组织线粒体功能和炎症状态从而改善肥胖,而SIRT1在其中起重要作用。本研究仅探讨了IF对白色脂肪组织的作用,需进一步检测其在组织代谢中的作用并通过体外实验更深入探讨SIRT1在其中的作用。