


材料与方法
一、材料及主要试剂
采用AEG-1 siRNA病毒载体,含绿色荧光蛋白(GFP)基因(psPAX,pMD2.G慢病毒包装系统,Invitrogen公司);结肠癌HCT116细胞株(购自中国模式培养物集库存,中国北京);免疫缺陷小鼠(购自中山大学实验动物中心)。Trizol 试剂(Invitrogen公司);RevertAid First Strand cDNA Synthesis Kit (#K1621,#1622,Fermentas公司)、SYBR Premix Ex Taq II (Tli RNaseH Plus) (DRR820)购自TaKaRa公司;RIPA 细胞裂解液和蛋白酶抑制剂(PMSF,广州博彩生物公司);Novex ECL 发光试剂盒(WP20005,Invitrogen公司);兔抗人AEG-1抗体;兔抗人GAPDH抗体(Invitrogen公司)。
二、方法
1.慢病毒介导干扰AEG-1-siRNA稳定细胞株构建
采用化学合成方法合成以AEG-1为靶点的RNA干扰序列(AEG-1-RNAi1,s:GCAGCAAGGCAGTCTTTAAGT,as:ACTTAAAGACTGCCTTGCTGC),挑选了带GFP报告基因的慢病毒载体(RNAil组)和空载体(空载组),采用psPAX,pMD2.G慢病毒包装系统,转染成功后收取上清(病毒液),病毒感染结肠癌细胞株HCT116(HCT116对照组),转染48 h后,采用0.5 μg/ml嘌呤霉素的10%胎牛血清DMEM培养基筛选出阳性细胞。
2. 实时荧光定量PCR(qRT-PCR)检测AEG-1mRNA的表达
采用Trizol试剂提取细胞的总RNA,以Oligo dT为引物逆转录成cDNA,逆转录条件为:65℃ 5 min,42℃ 60 min,70℃ 5 min。在ABI PRISM 7500 qRT-PCR仪上按照两步法进行扩增,使用β-actin作为内参,计算△CT值(△CT = CT目的-CT内参)和2﹣△CT。
3. 蛋白质印迹法检测AEG-1蛋白表达
采用RIPA裂解细胞,收集上清,定量,取50 μg总蛋白进行SDS-PAGE电泳,采用PVDF膜转膜,5%脱脂奶粉室温下封闭1 h后,分别加入兔抗人AEG-1抗体(Invitrogen) 1∶1000和兔抗人GAPDH抗体1∶1000,4℃过夜,TBST洗涤,加入相对应的二抗,室温孵育1 h,TBST洗涤,ECL显色试剂盒显影,以GAPDH为内参。
4. 裸鼠体内成瘤试验
将14只裸鼠随机分成2组即HCT116-control组和HCT116-RNAi1组,分别采用转染前后细胞株接种,将浓度为1×106个/ml的细胞悬液注射到裸鼠背部,每只0.2 ml,同样条件下饲养,每周测量接种部位瘤体的最长直径a(mm)和最短直径b(mm),瘤体体积= a×b2/2(mm3)。4周后处死小鼠取出瘤体测量质量。
三、统计学处理
采用SPSS 13.0进行统计分析。正态分布数据以$\bar{x}±s$表示,2组间比较采用t检验或者t'检验,重复测量资料采用重复测量资料方差分析,交互效应有意义时采用t检验分析单独效应,α= 0.05,两两比较采用Bonferroni法校正检验水准。
结果
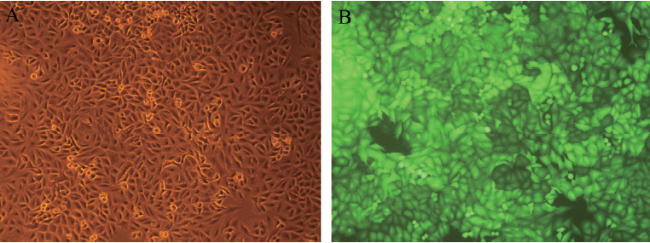
一、成功构建AEG-1-RNAi结直肠癌稳定细胞株
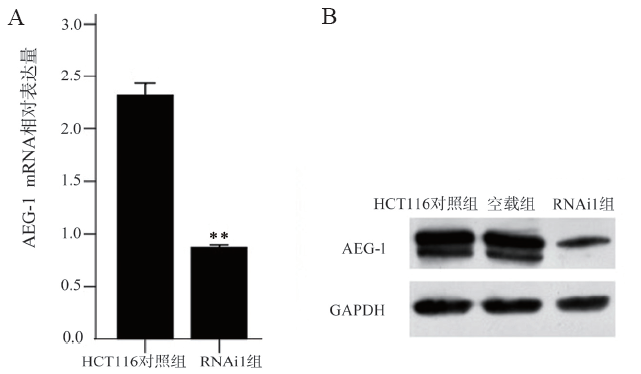
二、qRT-PCR和蛋白免疫印迹法检测AEG-1的干扰效果
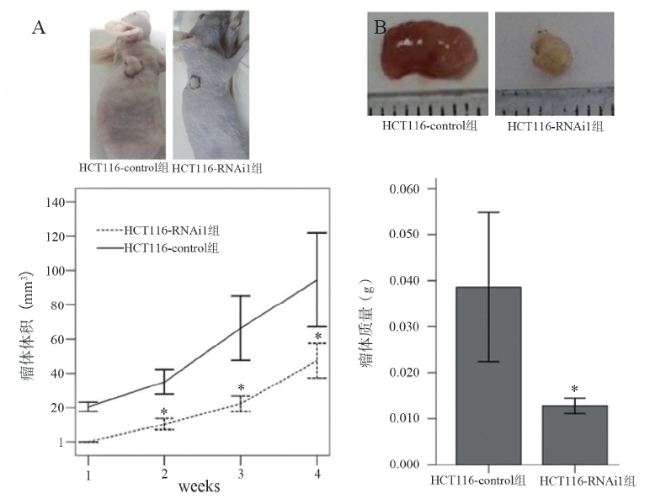
三、AEG-1对体内结肠癌细胞肿瘤发生的影响
第1周HCT116对照组处理的裸鼠已有明显的肿瘤形成(20.41±2.63)mm3,而注射AEG-1-RNAi处理的小鼠第1周肿瘤没有生长,随后2、3、4周瘤体体积分别为(10.43±3.30)、(22.31± 4.55)、(47.54±10.13)mm3,对照组为(35.14± 7.29)、(66.39±18.86)、(94.53±27.31)mm3(图3A),重复测量资料方差分析显示2组瘤体体积随时间变化的趋势不同,差异有统计学意义(F = 3.714,P = 0.025),因此分析单独效应。2组第2 (t = -6.906,P = 0.001)、3 (t = -5.080,P = 0.001)、4(t = -3.608,P = 0.007)周瘤体体积比较差异均有统计学意义。4周后处死裸鼠,取出瘤体测量质量,处理组为(0.013±0.002)g,低于对照组的(0.039± 0.016)g(t = -3.535,P = 0.023,图3B)。
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
讨论
大肠癌的演变是一个涉及多基因变化的复杂的过程,由正常肠上皮转化为肠上皮化生、腺瘤、腺瘤伴不典型增生、上皮内瘤变、癌,整个过程可能与多个癌基因、抑癌基因以及多条肿瘤信号通路的变化有关。近年来发现AEG-1不仅是各器官和组织致癌过程中的关键因素,而且是复杂的肿瘤信号通路网中的关键交汇点,其可以作用于多条信号通路,包括激活核因子-κB、Ha-Ras和磷脂酰肌醇3激酶/蛋白激酶B (Akt)通路、ERK/MAPK和Wnt/β-catenin通路,还有Aurora-A kinas信号通路等[12,13]。在非小细胞肺癌中,AEG-1通过提高抗凋亡蛋白Bcl-2水平和激活PI3K/Akt途径抑制细胞凋亡[9]。在结直肠癌中,AEG-1通过影响β-catenin增强结直肠癌细胞的转移和侵袭能力[13]。
我们前期研究以AEG-1基因为靶点,采用siRNA可以有效抑制AEG-1的表达,通过MTT、平板克隆、transwell和流式细胞仪等方法发现在体外下调AEG-1表达会抑制结肠癌细胞的增殖、克隆形成和侵袭能力,导致细胞周期G0/G1期停滞,促进凋亡和增加5-FU细胞毒性,以及影响血管生成相关因子缺氧诱导因子-1α(HIF-1α)的产生[5-6, 14]。然而,干扰AEG-1的表达在体内试验会有怎样的结果及其相关的信号通路,我们仍然不清楚。因此我们深入研究,进一步进行裸鼠体内成瘤实验。实验结果显示,沉默AEG-1可抑制结肠癌细胞在裸鼠体内成瘤的能力,抑制肿瘤的生长,可见AEG-1在结直肠癌的癌变过程中起着重要的作用,有望成为肿瘤基因治疗的新靶点和预后评估指标。
恶性肿瘤具有无限生长潜力,与它可以诱导血管生成是密切相关的,而AEG-1可以诱导新生血管生成及促进血管生成相关因子的产生。有学者报道AEG-1肝细胞的条件培养基可以诱导显著的血管生成;在宫颈癌中沉默AEG-1可以显著下调血管生成相关因子HIF-1α、Tie2、血管内皮生长因子(VEGF)和TEM1/CD248的表达,而且人脐静脉内皮细胞在SiHa细胞或转染AEG-1的正常宫颈上皮细胞的条件培养基下培养,其血管生成能力增强[15,16]。另也有报道AEG-1与微小RNA(miRNA)关系密切,AEG-1通过介导miRNA-154表达抑制胃癌细胞的增殖,miRNA-564直接作用于AEG-1抑制甲状腺乳头状癌的侵袭性,miRNA-504通过作用于AEG-1抑制视网膜母细胞瘤细胞的增殖和侵袭能力[17,18,19]。我们前期研究发现通过上调或下调AEG-1表达会改变miRNA的表达谱,推测AEG-1可能是通过改变某些miRNA的表达,从而影响血管生成相关因子的产生[5, 14]。前期研究显示上调AEG-1的表达会抑制miRNA-34a和促进HIF-1α的表达[14]。miRNA-34a是一个抑癌基因,高表达的miRNA-34a可抑制沉默信息调节因子1、损害内皮组细胞以及阻断VEGF的产物,从而抑制血管新生;而HIF-1α能使细胞更容易适应低氧的环境,是细胞对缺氧反应的最重要调节者,可以促进癌细胞表达VEGF,促进血管生成。有研究发现敲除miRNA-34a可通过激活Notch/HIF-1α信号通路,从而显著提高肝切除后10 d大鼠肝功能和肝细胞再生能力[20]。也有报道指出miRNA-34a通过负性调节SIRT1/HIF-1α信号来影响2型糖尿病相关耳蜗毛细胞的凋亡[21]。这与我们在结直肠癌中发现下调AEG-1的表达会促进miRNA-34a及抑制HIF-1α表达是一致的。因此我们推测在结直肠癌中,AEG-1可能通过抑制miRNA-34a促进HIF-1α的表达,从而促进VEGF的表达,发挥促进肿瘤血管新生的作用。
综上所述,沉默AEG-1可以抑制结直肠癌细胞的体内成瘤能力,其机制可能与通过调节miRNA从而影响血管生成相关因子的产生有关,这将为进一步研究结直肠癌中AEG-1基因治疗奠定基础。