


皮脂腺癌(SC)是一种罕见的、具有侵袭性的皮肤恶性肿瘤,好发于老年人,常表现为红色结节或者斑块,有时可出现溃疡,偶尔呈淡黄色。据报道,SC的常见部位为眼睑、面部、头皮等处。按生长部位,SC可分为眼周的SC和眼外的SC,其中眼外SC有不足2%发生于腮腺[1]。1953年,Foote和Frazell在一篇唾液腺肿瘤综述中首次报道了一例皮脂腺腺瘤(SA)。6年后,Rauch和Masshoff报道了腮腺中的SC。截至2022年12月,外文文献共报道了35例腮腺SC和10例腮腺SA[2-3]。由于其罕见性,腮腺SC的临床病理特征和组织发生尚未完全阐明,而对于相关基因突变目前国内外报道较少。本文对一例腮腺SC进行了肿瘤的全外显子组测序,由于腮腺SA更加罕见,本研究选择了同为眼外皮脂腺良性肿瘤的头皮SA样本作为对照,检测全外显子组基因突变在两者中的差异变化,从而初步探讨腮腺SC发生的可能机制及治疗的靶点。
材料与方法
一、标本来源
病例1 患者女,89岁。因发现耳后肿物2年于2022年9月至中山大学附属第三医院门诊就诊。彩色多普勒超声(彩超)检查提示右侧腮腺内实性肿物,肿物大小约4.0 cm×4.0 cm×3.5 cm,考虑恶性肿瘤性病变。手术切除肿物后行病理学活组织检查(活检),诊断为腮腺SC。
病例2 患者女,51岁。因发现头皮肿物5年于2020年6月至中山大学附属第三医院门诊就诊。通过详细有全身检查,排除头皮外其他部位的肿瘤。体格检查可见头皮隆起型肿物,肿物大小约2.5 cm×2.0 cm×1.5 cm,边界欠清,质硬。手术切除肿物后行活检,诊断为SA。
本研究方案经中山大学附属第三医院伦理委员会批准(批件号:中大附三医伦RG2023-117-01),患者均已签署知情同意书。
二、方法
1. 取样
切取石蜡标本中的肿瘤及瘤旁组织或瘤旁组织,体积约0.3 cm×0.3 cm×0.3 cm。
2. DNA的提取及建库
提取石蜡包埋组织的DNA,用Qubit对DNA浓度进行定量,以OD值在1.8~2.0,DNA含量>1.2.3 1.0 μg为建库的标准。将符合标准的DNA进行建库,具体操作同文献[4] 。
3. 数据处理及分析
筛选用于后续信息分析的高质量读序(clean reads),使用BWA软件将clean reads比对至人的参考基因组上,并通过Samtools软件处理,得到bam文件,再用picard软件去除PCR导致的重复reads,然后使用GATK进行矫正;之后基于比对结果,使用基因组分析工具箱(GATK)软件检测单核苷酸位点变异(SNV)及长度小于50 bp的插入和缺失(InDel),最后使用ANNOVAR软件对突变结果进行详细注释。
4. 筛选高频基因和驱动基因
5. 肿瘤异质性及亚克隆分析
利用SciClone软件将2例肿瘤标本的变异等位基因频率(VAF)及拷贝数变异(CNV)的拷贝数作为输入文件,通过识别体细胞突变的数量及分析亚克隆的组成,追踪亚克隆进化,得到每例肿瘤样本的克隆性图谱信息。
结果
一、病理结果
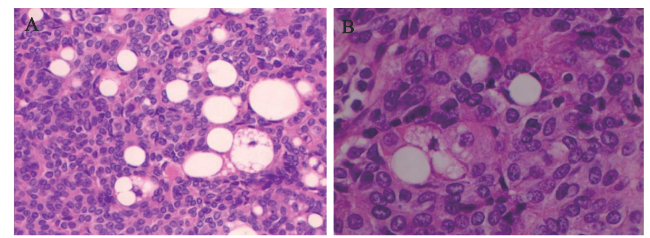
病例1 肿瘤呈浸润性生长,肿瘤细胞排列成大的巢团状,部分巢团中央可见坏死,巢团周边肿瘤细胞多呈基底细胞样,核质比高,细胞异型性明显,中央见散在体积较大、胞质呈多空泡状的皮脂腺分化细胞,符合SC。见图1。
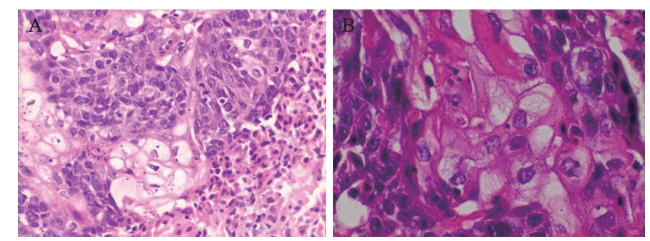
病例2 肿瘤边界清楚,由基底样细胞和成熟的皮脂腺细胞构成,基底样细胞大小形态较一致,细胞核呈卵圆形,可见小核仁,成熟皮脂腺细胞胞质丰富、呈多空泡状,细胞异型性不明显,未见确切核分裂及坏死,符合SA。见图2。
二、全外显子组测序及生物信息学分析结果
腮腺SC及其癌旁组织、头皮SA及其瘤旁组织的测序数据与参考基因组比对的结果均达到要求,与参考基因组比对能达到 94%以上的比对率;平均100倍测序深度时10倍以上覆盖度达到95%以上,见表1。腮腺SC组织的SNV数目为2012,头皮SA组织的SNV数目为205,腮腺SC组织的SNV数目约为头皮SA组织的10倍。腮腺SC组织中的SNV仅新发突变(未注释上千人基因组或dbSNP数据库)数目为1 495(占75%),头皮SA新发突变数目仅178(占86%),见表2。腮腺SC组织的SNV类型(图3)多于头皮SA组织(检测结果见文献[5] )。腮腺SC组织、头皮SA的InDel数目分别为2017、50,新发突变数目为1 070、50。而CNV的结果则分别为92和90。
表1 全外显子测序结果数据比对统计情况总览 |
| 样品名 | Clean reads/条 | 比对上参考基因组的reads/条 | 比对 率/% | 重复 reads/条 | 重复rates 比例/% | 探针捕获效率/% | 目标区域的平均测序深度/% | 目标区域的覆盖 度/% | 目标区域深度≥10倍的覆盖区域比例/% |
|---|---|---|---|---|---|---|---|---|---|
| 腮腺SC组织 | 175 543 260 | 165 903 139 | 94.51 | 48 313 747 | 29.12 | 75.57 | 210.68 | 97.17 | 96.29 |
| 腮腺SC癌旁组织 | 141 775 014 | 134 174 396 | 94.64 | 32 465 710 | 24.20 | 74.39 | 179.91 | 97.16 | 96.33 |
| 头皮SA组织 | 136 541 998 | 129 113 256 | 94.56 | 31 182 127 | 24.15 | 77.50 | 177.78 | 97.09 | 96.02 |
| 头皮SA瘤旁组织 | 102 327 688 | 96 319 832 | 94.13 | 22 671 397 | 23.54 | 77.65 | 133.71 | 97.10 | 95.88 |
表2 驱动基因突变筛选结果 |
| 基因名称 | 染色体 | 染色体上的位置 | 参考 碱基 | 突变碱基 | 突变功能 | CGC数据库中对应癌种 | 驱动基因 |
|---|---|---|---|---|---|---|---|
| ABL1 | chr9 | 1.34E+08 | A | G | 错义突变 | 慢性髓细胞性白血病、淋巴细胞白血病、T淋巴细胞-急性淋巴细胞白血病 | 高置信驱动基因 |
| ACTB | chr7 | 5568884 | AGA | — | 框内缺失突变 | — | 候选驱动基因 |
| ACVR2A | chr2 | 1.49E+08 | A | — | 移码缺失突变 | — | 候选驱动基因 |
| AHCTF1 | chr1 | 2.47E+08 | T | G | 错义突变 | — | 候选驱动基因 |
| ASPM | chr1 | 1.97E+08 | T | — | 移码缺失突变 | — | 高置信驱动基因 |
| ATF7IP | chr12 | 14633943 | C | A | 错义突变 | — | 候选驱动基因 |
| BAP1 | chr3 | 52441291 | — | T | 移码插入突变 | 葡萄膜黑色素瘤、乳腺癌、非小细胞肺癌、肾细胞癌 | 高置信驱动基因 |
| CDH1 | chr16 | 68835781 | C | — | 移码缺失突变 | 小叶乳腺癌;胃癌 | 高置信驱动基因 |
| CDKN2A | chr9 | 21974759 | C | — | 移码缺失突变 | 黑色素瘤、多种其他肿瘤类型 | 高置信驱动基因 |
| CHD4 | chr12 | 6707111 | C | T | 错义突变 | 子宫浆液性癌 | 高置信驱动基因 |
| CLIP1 | chr12 | 1.23E+08 | G | A | 错义突变 | 斯皮茨痣样黑色素瘤 | 高置信驱动基因 |
| CNOT1 | chr16 | 58577532 | — | T | 移码插入突变 | — | 候选驱动基因 |
| CNOT3 | chr19 | 54649671 | T | — | 移码缺失突变 | T淋巴细胞-急性淋巴细胞白血病 | 候选驱动基因 |
| COL18A1 | chr21 | 46924438 | CCCCCA | — | 框内缺失突变 | — | 高置信驱动基因 |
| COL18A1 | chr21 | 46876169 | C | T | 错义突变 | — | 高置信驱动基因 |
| CREB1 | chr2 | 2.08E+08 | G | T | 错义突变 | 透明细胞肉瘤、血管瘤样纤维组织细胞瘤 | 高置信驱动基因 |
| CYC1 | chr8 | 1.45E+08 | C | — | 移码缺失突变 | — | 候选驱动基因 |
| DDX3X | chrX | 41193874 | — | G | 移码插入突变 | 慢性淋巴细胞白血病、髓母细胞瘤 | 候选驱动基因 |
| DIDO1 | chr20 | 61510959 | G | A | 错义突变 | — | 候选驱动基因 |
| DMD | chrX | 32235050 | A | G | 错义突变 | — | 高置信驱动基因 |
| DNM3 | chr1 | 1.72E+08 | C | A | 错义突变 | — | 候选驱动基因 |
| DNM3 | chr1 | 1.72E+08 | C | T | 错义突变 | — | 候选驱动基因 |
| DST | chr6 | 56505257 | A | G | 错义突变 | — | 候选驱动基因 |
| EEF1A1 | chr6 | 74229650 | T | C | 错义突变 | — | 高置信驱动基因 |
| EEF1A1 | chr6 | 74228902 | G | A | 错义突变 | — | 高置信驱动基因 |
| FLNA | chrX | 1.54E+08 | T | C | 错义突变 | — | 候选驱动基因 |
| HUWE1 | chrX | 53577645 | C | — | 移码缺失突变 | — | 高置信驱动基因 |
| JAK2 | chr9 | 5066768 | T | — | 移码缺失突变 | 急性淋巴细胞白血病、急性髓系白血病、骨髓增殖性肿瘤、慢性髓细胞性白血病 | 高置信驱动基因 |
| LTBP1 | chr2 | 33525635 | G | A | 错义突变 | — | 候选驱动基因 |
| LZTR1 | chr22 | 21351194 | A | G | 错义突变 | 胶质母细胞瘤 | 高置信驱动基因 |
| MAST2 | chr1 | 46496749 | C | T | 无义突变 | — | 高置信驱动基因 |
| MDN1 | chr6 | 90399738 | A | G | 错义突变 | — | 候选驱动基因 |
| NCOA3 | chr20 | 46279887 | — | CAA | 框内插入突变 | — | 高置信驱动基因 |
| NCOR1 | chr17 | 15995261 | G | A | 无义突变 | 乳腺癌 | 高置信驱动基因 |
| NR4A2 | chr2 | 1.57E+08 | T | — | 移码缺失突变 | — | 高置信驱动基因 |
| NUP98 | chr11 | 3726447 | G | A | 错义突变 | 急性髓系白血病 | 高置信驱动基因 |
| P4HB | chr17 | 79817126 | C | T | 错义突变 | — | 候选驱动基因 |
| PLEC | chr8 | 1.45E+08 | C | G | 错义突变 | — | 高置信驱动基因 |
| PLEC | chr8 | 1.45E+08 | C | T | 错义突变 | — | 高置信驱动基因 |
| PLEC | chr8 | 1.45E+08 | G | A | 错义突变 | — | 高置信驱动基因 |
| PPFIA4 | chr1 | 2.03E+08 | C | T | 错义突变 | — | 候选驱动基因 |
| RAG1 | chr11 | 36596125 | AAG | — | 框内缺失突变 | — | 候选驱动基因 |
| RNF43 | chr17 | 56435161 | C | — | 移码缺失突变 | 胆管癌、卵巢癌、胰腺癌 | 高置信驱动基因 |
| RNF43 | chr17 | 56435193 | AG | — | 移码缺失突变 | 胆管癌、卵巢癌、胰腺癌 | 高置信驱动基因 |
| RPL22 | chr1 | 6257785 | T | — | 移码缺失突变 | 急性髓系白血病、慢性髓细胞性白血病 | 高置信驱动基因 |
| RXRA | chr9 | 1.37E+08 | G | A | 错义突变 | — | 高置信驱动基因 |
| SETD2 | chr3 | 47164922 | G | A | 错义突变 | 肾透明细胞癌 | 高置信驱动基因 |
| SPTAN1 | chr9 | 1.31E+08 | T | C | 错义突变 | — | 候选驱动基因 |
| SRRM2 | chr16 | 2816326 | C | T | 错义突变 | — | 候选驱动基因 |
| STAT3 | chr17 | 40491387 | G | A | 错义突变 | T细胞大颗粒淋巴细胞白血病 | 高置信驱动基因 |
| TBL1XR1 | chr3 | 1.77E+08 | T | — | 移码缺失突变 | 脾边缘区淋巴瘤、原发性中枢神经系统淋巴瘤、结直肠癌、胆囊癌 | 高置信驱动基因 |
| TCF7L2 | chr10 | 1.15E+08 | A | — | 移码缺失突变 | 结直肠癌 | 高置信驱动基因 |
| TET2 | chr4 | 1.06E+08 | A | T | 错义突变 | 骨髓增生异常综合征 | 高置信驱动基因 |
| TTK | chr6 | 80751897 | A | — | 移码缺失突变 | — | 候选驱动基因 |
| UHRF1BP1L | chr12 | 1E+08 | G | T | 错义突变 | — | 高置信驱动基因 |
| ZFHX3 | chr16 | 72993474 | A | G | 错义突变 | 子宫内膜癌、胃癌、前列腺癌 | 高置信驱动基因 |
| ZMYND8 | chr20 | 45849979 | T | A | 无义突变 | — | 候选驱动基因 |


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
讨论
目前有关眼外SC的基因检测分析数据极其罕见,尤其是腮腺SC的结果尚未见报道。本研究通过对腮腺SC和头皮SA进行全外显子测序分析,发现腮腺SC的基因突变总数和各突变类型均远远超过头皮SA,说明腮腺SC的基因突变非常活跃。此外,腮腺SC中发现有别于头皮SA的 57个驱动基因突变,其中比较明确的癌症相关基因有ABL1、ACTB、ASPM、ZFHX3和NCOA3基因等。ABL1基因是与白血病相关的原癌基因。当其易位到 22 号染色体长臂上时,会与断裂点丛集区(BCR)基因发生融合从而形成 BCR-ABL 融合基因,该基因编码的BCR-ABL融合蛋白持续性活化,导致了白血病[10]。ABL1激酶在实体瘤中的活化并不是因为发生染色体易位形成BCR-ABL融合蛋白,而是由于 ABL1 激酶表达上调或ABL1蛋白的上游激酶活化从而提高了 ABL1的酪氨酸激酶活性。ACTB被广泛用于量化肿瘤表达水平。然而,ACTB聚合与ACTB细胞骨架形成的驱动细胞突起和细胞运动参与了癌症的侵袭和转移[11]。ASPM基因位于人类1号染色体,ASPM在多种癌症中异常表达,且与肝癌、卵巢癌、肺腺癌、胰腺癌和膀胱癌等多种恶性肿瘤的不良预后相关[12⇓-14]。其缺失会影响细胞有丝分裂、分裂方向和分化方式,造成DNA损伤增加和细胞凋亡。ZFHX3作为非小细胞肺癌免疫检查点阻断的生物标志物,是已知的原癌基因,也是基底细胞癌相关基因[15-16]。NCOA3基因与癌症相关,其体细胞基因拷贝数发生变化可导致转录水平发生变化,从而导致乳腺癌和胰腺癌的发生、发展。
头颈部肿瘤的发展是一个综合作用的结果,多种基因的突变相互作用,相互促进,形成了恶性循环。肿瘤异质性即肿瘤组织内部不同的肿瘤细胞或亚群中体细胞突变不完全相同。大部分的肿瘤存在亚克隆,而亚克隆突变与耐药性相关。分析肿瘤的克隆结构有利于揭示肿瘤组织的异质性。分析肿瘤异质性和克隆结构有助于研究肿瘤的发展、进化、转移、复发以及药物反应等。本研究中,腮腺SC的肿瘤异质性较头皮SA高,亚克隆多。本研究在腮腺SC中发现了ABL1基因的突变,而针对ABL1突变发生的肿瘤,临床有潜在作用的治疗药物包括伊马替尼、达沙替尼、尼洛替尼、波苏替尼、泊那替尼等。
综上所述,腮腺SC具有不同于良性肿瘤的基因突变及突变的模式。但本研究样本量有限,进一步的研究需要更多的样本进行相关基因突变的验证。